Reproduction
des
Sarracenia
Homepage > Dossiers > Sarracenia > Reproduction
Mise à jour :
mardi 21 décembre, 2010
Révision complète le vendredi 23 mai 2008
© 2007 J.Ph. ROSELLO
Dans le genre Sarracenia il n'y a de possible que le partage du rhizome. Avant d'envisager quoi que ce soit, sachez que si vous divisez un plant qui ne produit pas encore d'urnes adultes, vous risquez ralentir leur apparition. Il y a tout de même diverses modalités plus ou moins adaptées aux espèces ou hybrides : Il s'agit de grandes lignes et cela dépend du clone, c'est-à-dire de la lignée. Des différences se voient même dès le semis. Les plantes qui forment peu de pousses ont intérêt à être partagées plus souvent pour augmenter leur nombre, sans quoi on se retrouve avec une masse de rhizome et trois feuilles au bout. Dans la nature c'est une sécurité qui peut garantir la survie, mais vous êtes là pour veiller sur votre plant... L'idéal serait de procéder au printemps au tout début de la reprise. Le pire est donc l'opposé, l'automne, au moment où la plante entre en repos et les blessures aux racines seront autant de portes d'entrée pour les moisissures. Voir Les rempotages. Un autre mode est la culture in vitro. Ses principes sont identiques pour les Sarracenia que pour la Dionée - voir le sujet. Remarques prélables Les fleurs des Sarraceniacées sont souvent intéressantes, voire très belles. Mais si la reproduction par graines (donc à partir de fleurs) est très difficile pour le genre Heliamphora, d'intérêt assez limité pour le genre Darlingtonia qui n'a qu'une espèce donc pas d'hybride, celle des Sarracenia est vraiment très facile à réaliser artificiellement et permet toutes les combinaisons. Les fleurs sont hermaphrodites (à la fois mâle et femelle) et apparaissent de la fin de l'hiver au milieu du printemps, avec un certain étalement selon l'espèce : S. flava est très précoce, S. minor est tardive, fleurit jusqu'en juin. Des fleurs peuvent apparaître très en retard mais avorteront. J'ai même eu une fleur fin septembre, pratiquement sans tige donc rester à l'envers et néanmoins parfaitement ouverte... Plutôt que de longues explications, voici tout de même comment on écrit les noms concernés, car on trouve de tout sur le Net :
Rappelons qu'il y a huit espèces du genre Sarracenia reconnues actuellement, SS. alata, flava, leucophylla, minor, oreophila, psittacina, purpurea et rubra, pas une de plus ! Mais S. flava présente plusieurs variétés, et d'autres espèces ont quelques sous-espèces. D'après les conventions botaniques actuelles, la forme qui sert de référence et que l'on espère (ou suppose) souvent comme l'origine du groupe est nommée actuelle "forme type" mais il y a peu on doublait tout simplement le nom. Par exemple la forme type de S. rubra est "S. rubra type", ce que l'on écrivait il y a quelques années comme "S. rubra rubra". Vous verrez encore cette écriture mais évitez-la. Le nom du type était donc traité comme une sous-espèce virtuelle qui portait le même nom. Ceci est valable aussi pour les variétés sauvages. On n'écrit donc plus S. flava flava ou S. flava var. flava mais S. flava type ou S. flava forme type. Pour S. rubra, il y a en plus de la forme type, ssp gulfensis, alabamensis, jonesii et wherryi ; pour S. purpurea, il y a la forme type et ssp. venosa avec quelques variations et pour S. minor il y a en plus ssp okefenokeensis, géante et aquatique qui est le Sarracenia qui fleurit le plus tard (chez moi en tout cas) sans doute à cause de l'eau qui maintient une température plus basse plus longtemps au printemps. Si vous étudiez un hybride, tenter de retrouver les parents est très incertain ! Pour un plant immature, vous pouvez retenir que la couleur des jeunes plants est souvent plus vive mais moins nuancées : les détails comme certaines taches ne se formeront pas ou très peu. Il y a heureusement des signes qui permettent tout de même de retrouver les espèces ancestrales mais c'est la proportion, le rang de chacun, qui est beaucoup plus difficile. Vous trouverez ces signes dans ma page sur l'hybridation des Sarracenia. Voici le dessin d'une fleur. Elle est d'ordre 5 donc tous les organes sont au nombre de 5 ou multiples, en tout cas dans la fleur parfaite. Il y a ainsi 5 sépales rigides et souvent colorés mais trois d'entre eux ont été enlevés, ainsi que beaucoup d'étamines.. Il y a aussi 5 pétales enlevés également pour montrer l'intérieur. Les 3 "sortes de sépales" tout en haut sont des restes de l'évolution et leur aspect est très variable, il s'agit en fait de bractées, feuilles spéciales très souvent modifiées qui accompagnent les fleurs mais n'en font pas partie. On voit qu'elles sont à l'écart de la fleur bien que souvent colorées elles aussi. 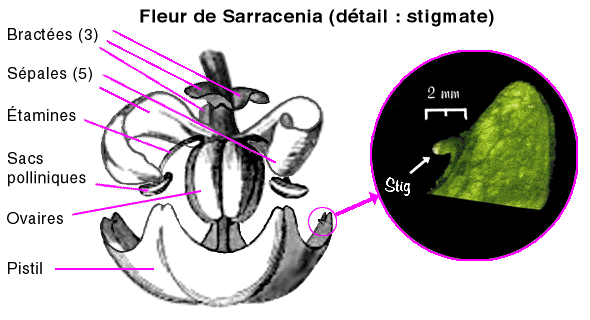 Technique d'obtention Les grains de pollen à maturité tombent en masse des étamines et arrivent dans le creux du "parapluie" que forme le pistil et il ne faut pas tarder à les utiliser.Il suffit de prélever le pollen et de le déposer sur les stigmates disposés à l'extrémité des "baleines" du "parapluie". Plutôt qu'un pinceau, j'utilise de préférence le petit doigt pour récupérer, déposer et presser ce pollen sur les stigmates, qui sont à peine assez collants pour retenir les grains : le pinceau en dépose peu, et la détente inopinée des poils en projette beaucoup dans l'air (même si c'est invisible), ce qui ne garantit plus la pureté des croisements alentours. De plus, s'il est prévu de récolter le pollen de cette fleur celui-ci sera pollué par le pollen étranger projeté. Lorsque tous les sacs polliniques ont largué leur pollen, la fleur n'est pratiquement plus ou très peu féconde (expérience personelle), à plus forte raison quand les pétales tombent. Pour un résultat optimal, la fleur mère devrait donc être à peine éclose. Pour un usage différé, le pollen doit être conservé au réfrigérateur (comme les graines) si possible avec un agent dessiccant (= qui absorbe l'humidité) : lorsque, par exemple, S. minor fleuri cela fait longtemps que les fleurs de S. flava sont fanées... En effet, si l'enveloppe protéique des grains de pollen a la réputation d'être "indestructible" (cf. pollen fossile) ceux-ci perdent leur fécondité assez rapidement.
Les pétales tombent rapidement et le reste se maintient très longtemps, souvent jusqu'à l'automne. C'est une manière de conserver un stock de graines loin du sol, qui pourra même germer plusieurs années après. L'évolution a sans doute conservé ce mécanisme parce que seules les plants qui présentaient cette particularité ont survécu à des sécheresses locales... La fécondation produit en général un effet intéressant : le redressement partiel ou total de la fleur. L'intérêt n'est pas évident... Je pense que cela permet au pollen accumulé dans "le parapluie" du pistil de tomber, se disperser au vent au lieu d'y pourrir aidé par l'eau qui s'y dépose. Le phénomène, me semble-t-il, accélére la chute des pétales : il n'est pas rare de les voir tomber dès le lendemain et cela permet peut-être aussi d'éviter quelques moisissures en évitant le confinement. Il est conseillé généralement dans la littérature de répéter plusieurs fois l'opération, une fois par jour pendant plusieurs jours, ce qui peut être délicat : si l'on veut être sûr de l'origine des parents l'utilisation d'un même pinceau pour des croisements variés nécessite lavage à l'eau courante, trempage dans l'alcool et séchage après chaque opération. En utilisant l'auriculaire, le nettoyage est simplifié, il suffit de le passer sur une éponge récurante pour décoller tous les grains. Il est vraiment regrettable de voir des plantes multipliées sans aucun soins par graines et porter des noms manifestement incorrects et, pire, être diffusés sous ce nom. Pour reproduire une espèce afin de limiter l'autofécondation, il vaut mieux utiliser des plants de sources différentes (le terme exact est clones différents) car il y a en effet un risque de dégénérescence. Cela n'a rien d'obligatoire. Lors d'hybridation, il arrive spontanément que du pollen d'un pied féconde une de ses propres fleurs, vous aurez du mal à l'éviter. Soyez donc prudent lorsque vous faites des semis : si le plant ne présente aucun caractère du (donc le donneur de pollen) vous avez au mieux une autofécondation. Cela ne veut pas dire non plus que ceux-ci seront identiques au pied : des gènes récessifs se manifesteront car des gènes dominants disparus n'auront plus leur effet de "masquage", etc. Est-il besoin de dire que si vous croisez des plants issus de la division d'un même pied, il s'agit tout de même d'autofécondation ? Les gènes restent, bien sûr, les mêmes chez les deux parents...
Les fruits et graines Au cours de l'été, l'ovaire se transforme en fruit (au sens botanique du terme...) et contient à maturité, à la fin de l'été et en début d'automne, des centaines de graines comme le montre cette "photo" (objets directement scannés). Dans un seul fruit de S. purpurea venosa, j'ai compté jusqu'à 800 graines. Il faut compter quatre mois de maturation. Leur couleur varie avec l'espèce du pied porteur, donc la mère : brun café, brun chocolat, brun rosé (S. minor)... Mais on observe beaucoup de différences selon les fruits d'une même plante. Le fruit peut rester assez petit et être plein de graines comme beaucoup grossir et être plein d'air ! C'est assez bluffant et cela rrive en particulier sur ceux qui sont allongés. Les fruits bien ronds et gros ne trompent pas, on dirait qu'ils vont éclater et lorsqu'on tape dessus avec l'ongle, on obtient un bruit compact, mat. Inversement, S. purpurea produit des fruits gros et ronds mais souvent creux et sans la moindre graine.  Fruit ouvert et graines de Sarracenia minor Elles seront conservées au froid de préférence et semées au début du printemps de l'année suivante ou ultérieurement mais certains les sèment immédiatement, si elles sont obtenues assez tôt. Elles ne sont pas éternelles toutefois. Après semis, il n'est pas rare de voir quelques graines germer une ou plusieurs années plus tard. N'oublions pas que la tourbe conserve bien et c'est une sacrée sécurité pour des milieux humides souvent susceptibles de sécheresse exceptionnelle. Résultats Maintenant que je vous ai fait un peu rêver, il faut vous avouer que la première année les plants mesurent environ 2,5 cm de haut, puis 5 cm. Ils sont alors peu colorés, bien que les tendances soient déjà marquées en général. Les pièges sont primitifs, dits "juvéniles" peu différents d'un croisement à l'autre, si ce n'est au niveau de leur inclinaison. Par exemple, les descendants des S. purpurea ou S. psittacina auront très tôt des feuilles obliques, un peu penchées vers l'extérieur... C'est le moment de rempoter car la troisième année, la taille est d'une vingtaine de centimètres et la plante prend une bonne partie de ses caractéristiques adultes, bien que "simplifiés". Elle fleurit la quatrième ou la cinquième année, ce qui vous permet de voir, enfin, les caractères de la fleur, de réaliser un back-cross avec l'un des parents, etc.
Les jeunes plants ont souvent des urnes qui poussent d'abord couchées ("prostrées") alors que les adultes (à ce stade les parents et grands-parents) n'ont pas ce caractère. On pourrait considérer que cela pourrait présenter un avantage pour des plants minuscules mais, n'en voyant aucun, il peut s'agir aussi d'un caractère ancestral. Il en résulte (ce n'est que mon avis) que l'ancêtre des Sarracenia avait sans doute seulement ce type d'urne et peut-être même qu'il était très proche de S. purpurea. Ce phénomène d'urnes couchées existe aussi dans le genre Darlongtonia mais non le genre Heliamphora. On remonte peut-être à l'ancêtre commun des Sarracenia et des Darlingtonia...  Cette Sarracenia flava fortement rayée est sans doute une création horticole : la véritable forme "fortement veinée", dont le qualificatif exact est ornata, présente à l'extrémité de la feuille un prolongement typique qui s'enroule sur lui-même (ce qui n'est pas le cas ici). Ce plant résulte vraisemblablement du croisement entre les variétés "ornata" et "rugelii". Ce n'est donc pas un hybride au sens strict (même espèce). Cette Sarracenia flava fortement rayée est sans doute une création horticole : la véritable forme "fortement veinée", dont le qualificatif exact est ornata, présente à l'extrémité de la feuille un prolongement typique qui s'enroule sur lui-même (ce qui n'est pas le cas ici). Ce plant résulte vraisemblablement du croisement entre les variétés "ornata" et "rugelii". Ce n'est donc pas un hybride au sens strict (même espèce).Le sélectionneur a conservé parmi la descendance le plant qui a hérité des qualités intéressantes des parents. |
