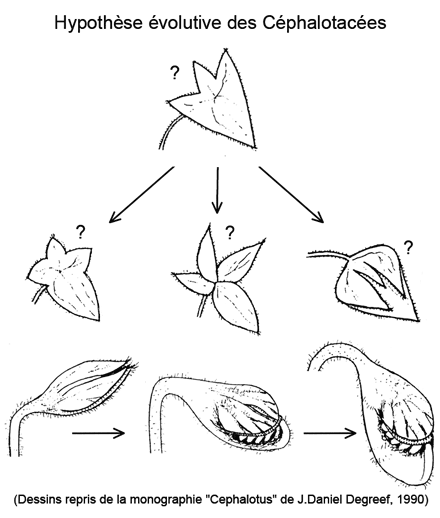
Hypothèse sur l'évolution du Cephalotus
Degreef, dans sa monographie nommée "Cephalotus" (1990) nous explique que l'étude des faisceaux conducteurs de la sève révèle la présence de quatre lobes originels. En gros, trois d'entre eux, plus ou moins séparés, plus ou moins repliés, se seraient soudés pour former le couvercle du piège. Le quatrième lobe se serait développé à l'extrémité de la feuille, incurvé, prenant une forme évasée qui conduit à l'urne proprement dite.
Il nous présente une hypothèse sur la manière dont ce serait formé ce curieux piège, qui n'apparaît qu'en été, les feuilles d'hiver ayant un aspect banal, simple :
D'après ses dessins, on ne comprend pas très bien quelle fonction pourrait bien remplir les stades intermédiaires repliés en forme de bouton...
L'origine de la feuille d'hiver (donc sans piège) est plus surprenante. Les coupes au microscope montrent cette fois que qu'elle correspond seulement au lobe central du couvercle et non à la feuille entière, banale. Degreef en conclue que la feuille simple est un piège avorté.
Cette conception est pour moi une approche qui reste un peu trop centrée sur le piège. Au sens strict, si la feuille d'été n'était qu'un piège avorté il ne serait qu'un bout de couvercle très fin. Or, il s'agit d'une feuille très épaisse (dite "succulente" car pleine de "suc") large, riche en éléments de réserve, beaucoup moins fragile que le piège. Elle est tout aussi importante pour la plante, même si ce n'est pas la même chose pour nous ;-). Ce n'est qu'un détail, l'hypothèse reste inchangée, mais je préfère voir une même ressource embryologique qui, au fil du temps, a conservé deux voies possibles tout aussi utiles :
- développement du lobe inférieur qui se "creuse" en coupelle, orientant une croissance des trois lobes supérieurs opposés vers une fusion et la formation d'un couvercle membraneux, translucide.
- ou, au contraire, développement d'un lobe supérieur qui inhibe complètement le développement des autres, concentrant ainsi les ressources pour former une feuille épaisse, riche en réserves, qui pourra plus facilement régénérer une plante entière si elle se casse ou si le rhizome meurt.
Il faut donc imaginer une structure intermédiaire entre la feuille originelle banale (mais tout de même composée de lobes) et le stade actuel. Une telle structure n'est plausible que si elle est fonctionnelle : cette feuille disparue inconnue devait donc rester efficace vis-à-vis de la photosynthèse, du stockage de réserves, tout en permettant de récolter un peu d'eau et permettre déjà la noyade d'insectes. Ce n'est pas le cas de l'hypothèse représentée. C'est cette noyade qui a été décisive car la décomposition spontanée de l'animal va libérer de précieux sels minéraux. Cette feuille encore primitive apporte déjà un avantage évolutif considérable par rapport aux feuilles ancestrales.
Le piège actuel est également capable de sécréter de l'eau : il est devenu indépendant de la pluie. Dès que cette capacité est apparue autrefois, ce qui n'est pas une évolution révolutionnaire, les trois lobes supérieurs ont pu varier car il n'était plus nécessaire qu'ils laissent libre l'ouverture pour la pluie. Ils ont pu notamment s'incurver vers le lobe inférieur en coupelle, ce qui permet un deuxième avantage évolutif : l'urne ne risque plus de déborder en cas de fortes précipitations et les sels minéraux ne sont donc plus dilués et perdus. Les lobes supérieurs ne servent plus que comme couvercle et peuvent même devenir translucides, leurre classique pour les insectes qui essaient de s'échapper et s'épuisent.
On remarquera que, de cette époque, vient certainement la capacité du couvercle à se refermer lorsque l'air est sec et se soulever dans le cas contraire. Ce mécanisme n'est pas anodin.
Puisqu'il n'y a plus dilution, les sécrétions d'eau à l'intérieur de l'urne peuvent aussi permettre la libération de substances qui existent dans toute cellule et qui servent à détruire les "pièces usées" de la cellule pour les recycler. Ces substances "corrosives" ne sont pas libres dans la cellule, heureusement, mais enfermées dans des vésicules et ce sont celles-ci qui finissent pas englober les organites à dissoudre. Il s'agit tout simplement d'enzymes. Ces vésicules sont mobiles et dès qu'elles ont pu se vider à l'extérieur de la cellule, en l'occurrence dans l'urne, les enzymes ont été détournés de leur rôle initial pour servir tout simplement d'enzymes digestifs.
Ce piège encore simple n'a, dès lors, plus cessé d'évoluer dans sa fonction : les parois intérieures ont produit les deux types de cellules actuelles spécialisées dans la digestion (cas unique) et d'autres cellules, au lieu de sécréter de l'eau ou des enzymes, ont libéré de plus en plus de sucres d'où la formation de glandes nectarifères. Ces glandes sont d'autant plus efficaces qu'elles sont difficiles d'accès, odorantes et protégées de l'eau : il y a donc encore un avantage évolutif qu'elles se soient développées plutôt sur le bord de la coupelle mais protégées par le replis de la feuille vers l'intérieur. Ce rebord est donc plus efficace s'il est légèrement découpé, d'où la formation de dents, d'abord modestes, de plus en plus susceptibles d'empêcher les insectes de s'échapper, encore un avantage...
Finalement, le plus curieux finit par être l'existence des feuilles simples, surtout avec cette origine embryologique ! Après tout, c'est le seul cas parmi les plantes carnivores et l'urne d'autres espèces remplit très bien son rôle au niveau de la photosynthèse. Comment expliquer cette particularité ?
Il y a une différence jamais évoquée entre les deux types de feuilles : le piège est fortement velu, les poils jouant un rôle notamment de guidage des insectes vers l'orifice, effet renforcé par les cannelures à l'extérieur. Le couvercle est velu sur les bord mais pas au centre, zone qui, on l'a vu, est l'origine embryologique de la feuille simple, la seule partie de la plante parfaitement lisse, absolument dépourvue de poils. Si vous cultivez cette plante vous avez sans doute remarqué que les poils très raides du Cephalotus ont la fâcheuse tendance à retenir fortement toutes les saletés, les éclaboussures, les restes divers : la plante vit en sous-bois et tous les déchets retombant sont préjudiciables à la photosynthèse. L'idée peut donc sembler étrange mais pour moi ce développement du lobe supérieur central, au détriment des autres, pour former finalement une feuille bien lisse est très favorable à la plante, tout au moins quand il y a peu d'insectes et peu de lumière.
Pendant la belle saison, les insectes abondent, la lumière devient suffisante, le lobe inférieur des feuilles en formation n'est plus inhibé, il draine les ressources au détriment des lobes supérieurs qui vont rester minces et plus petits. Tous les lobes restant longtemps soudés, l'ensemble va se gonfler d'air, grossir, essentiellement au niveau du lobe inférieur. La paroi du piège est plus épais sur l'avant, sinon cette espèce de bulle serait très fragile. Cet épaississement a pu évoluer à son tour pour former cette sorte de rail velu qui empêche les insectes de passer latéralement et les oblige à monter vers l'entrée du piège. Voilà encore une évolution bénéfique !
Au moment de l'ouverture les trois lobes supérieurs restent soudés en couvercle et le lobe inférieur est creusé en urne. Les tissus vont se renforcer, le couvercle va prendre des couleurs vives qui le font confondre avec une fleur, encore un petit plus pour l'attraction...
Caltha dionaeifolia
Une curieuse plante existe en Terre de Feu. Cette plante y est assez abondante par endroit et pousse dans les tourbières à Sphaignes, à l'extrême pointe de l'Amérique du Sud... Elle n'est pas carnivore mais présente des feuilles particulières dont les deux lobes internes ressemblent à des pièges de Dionée. Ces feuilles auraient la capacité de se refermer la nuit, peut-être pour se protéger du froid... Ce mécanisme est assez banal mais l'aspect général de la plante est vraiment unique.
Le Pr. Jolivet a présenté la plante en 1989 à des amateurs de plantes carnivores dont J.Daniel Degreef, qui en a réalisé deux schémas visibles dans sa monographie sur le Cephalotus :
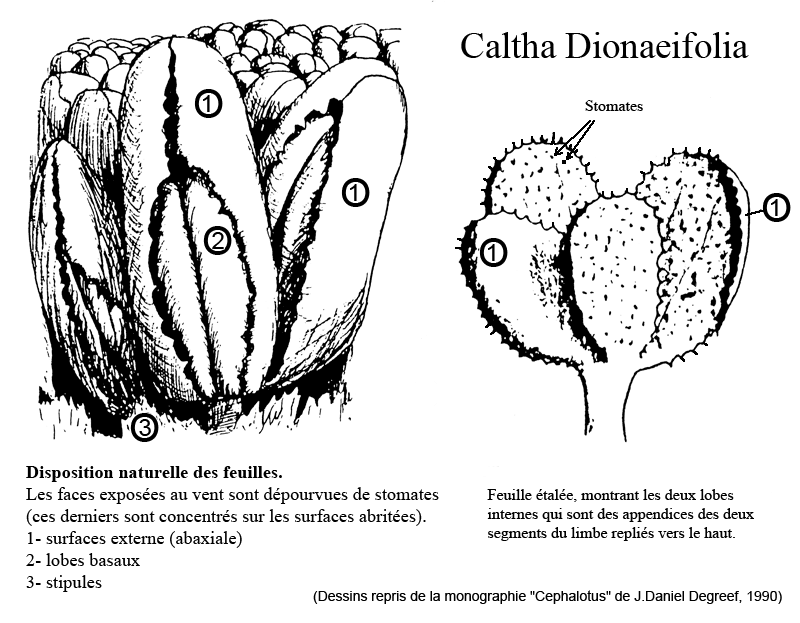
Le nom le plus fréquent semble être "dionaeifolia" bien que la déclinaison latine devrait être "dionae-ae" (comme ros-ae) donc le mot "dionaeaefolia" c'est-à-dire "feuille de Dionée", mais on trouve aussi "dionaefolia", "dionefolia", "dioneifolia"... La plante est très peu connue au point qu'aucune photo de la plante vivante n'est visible sur le Net.
A l'adresse http://www.botany.utoronto.ca/courses/bot299y/Caltdion.html on peut trouver le document ci-dessous.
Si vous avez mieux, je suis preneur ! Je suis à la recherche d'autres informations sur cette plante voire la plante elle-même.
Je me suis permis de traiter la photo, de très mauvaise qualité à l'origine. Voici le résultat :

 Le nom des plantes cultivées Le nom des plantes cultivées
Les amateurs utilisent utilisent et recopies de nombreuses variations autour des noms de leurs plantes. Voici un chapitre pour y voir plus clair.
Le première source est la traduction de l'anglais du Code international de nomenclature botanique, de 1999 (St Louis). La partie qui nous intéresse est l'article 28, situé dans le chapitre III de la section 6. Comme il y a plusieurs renvois, voici l'article avec des hyper-liens vers une copie des renvois un peu plus bas, Web oblige ;-). J'ai ajouté quelques couleurs et deux virgules (note 5) sans lesquelles le texte serait incompréhensible :
Les plantes sauvages introduites en culture gardent les noms qui sont appliqués aux mêmes taxons croissant dans le milieu naturel.
Note 1. Les hybrides, y compris ceux apparus en culture, peuvent recevoir des noms conformément aux dispositions de l’App. 1 (voir aussi les Art. 11.8 [Règles de priorité], 40 [Nom latin d'un hybride], et 50 [Recommandations générales relatives aux citations]).
Note 2. Les désignations indépendantes, supplémentaires pour des catégories spéciales de plantes employées dans
l’agriculture, sylviculture, et horticulture (et survenant que ce soit dans la nature ou en culture) sont traitées avec dans le Code International de Nomenclature pour les Plantes Cultivées, où le terme « cultivar » est défini et les règles sont fournies pour la formation et l’emploi des épithètes de cultivar.
Note 3. Rien n’écarte l’emploi, pour les plantes cultivées, de noms publiés conformément aux exigences du Code
botanique.
Note 4. Les épithètes dans les noms publiées en conformité avec le Code botanique peuvent être employées comme
des épithètes de cultivar selon les règles du Code International de Nomenclature pour les Plantes Cultivées, quand
le cultivar est considéré être le statut approprié pour les groupes concernés.
Ex. 1. Mahonia japonica DC. (1821) peut-être traité comme un cultivar, qui est alors désigné comme Mahonia ‘Japonica’ ; Taxus baccata var. variegata Weston (1770), quand il est traité comme un cultivar, est désigné comme Taxus baccata ‘Variegata’.
Note 5. Le Code International de Nomenclature pour les Plantes Cultivées fournit, pour l’établissement, des épithètes
de cultivar différant sensiblement d’épithètes de forme latine.
Ex. 2. ×Disophyllum ‘Frühlingsreigen’. Eriobotrya japonica ‘Golden Ziad’et E. japonica ‘Maamora Golden
Yellow’; Phlox drummondii ‘Sternenzauber’ ; Quercus frainetto ‘Hungarian Crown’
Ex. 3. Juniperus ×pfitzeriana ‘Wilhelm Pfitzer’(P. A. Schmidt 1998) était établi pour un cultivar présumé
tetraploide résultant du croisement original entre J. chinensis L. et J. sabina L. |
Article 11.8. Aux fins de la priorité, les noms latins donnés aux hybrides suivent les mêmes règles que ceux des taxons non hybrides de rang correspondant.
Ex. 30. Le nom ×Solidaster H. R. Wehrh. (1932) a priorité sur ×Asterago Everett (1937) pour l’hybride entre Aster L. et Solidago L.
Ex. 31. Anemone ×hybrida Paxton (1848) a priorité sur A. ×elegans Decne. (1852), pro sp., comme binôme des
hybrides provenant d’A. hupehensis (Lemoine & E. Lemoine) Lemoine & E. Lemoine × A. vitifolia Buch.-Ham. ex DC.
Ex. 32. Camus (in Bull. Mus. Natl. Hist. Nat. (Paris) 33 : 538. 1927) a publié le nom ×Agroelymus A. Camus
comme nom d’un nothogenre mais sans diagnose ni description latine, mentionnant uniquement les noms des
parents (Agropyron Gaertn. et Elymus L.). Comme le nom n’était pas validement publié suivant le Code alors en
vigueur, Rousseau, (in Mém. Jard. Bot. Montréal 29 : 10-11. 1952), a publié une diagnose latine. Toutefois,
conformément au présent Code (Art. H.9), la date de publication de ×Agroelymus est 1927 et non 1952, et il a ainsi priorité sur le nom ×Elymopyrum Cugnac (in Bull. Soc. Hist. Nat. Ardennes 33 : 14. 1938). |
Article 40
40.1. Pour être validement publié le nom d’un hybride de rang spécifique ou infraspécifique ayant une épithète latine doit être conforme aux règles qui s’appliquent aux noms des taxons non-hybrides de même rang.
Ex. 1. « Nepeta ×faassenii » (Bergmans avec description néerlandaise (Vaste Pl. ed. 2. : 544. 1939) et Lawrence
avec description anglaise (Gentes Herb. 8 : 64. 1949), n’est pas validement publié, parce qu’aucune description ou
diagnose latine, ni aucune référence à une telle description ou diagnose, ne l’accompagne. Par contre, le nom Nepeta
×faassenii Bergmans ex Stearn (1950) est validement publié parce qu’une description latine l’accompagne.
Ex. 2. « Rheum ×cultorum » (Thorsrud & Reisaeter, Norske Plantenavr. : 95. 1948), étant ici un nomen nudum n’est
pas validement publié.
Ex. 3. « Fumaria ×salmonii » (Druce, List Brit. Pl. : 4. 1908) n’est pas validement publié, parce que l’auteur s’est
borné à indiquer les parents présumés de l’hybride, F. densiflora × F. officinalis.
Note 1. Pour les noms d’hybrides au rang de genre ou de subdivision de genre, voir l’Art. H.9. |
Recommandations générales relatives aux citations
Recommandation 50A
50A.1. La citation d’un nom publié comme synonyme et donc invalide devrait être suivie de l’expression « comme
synonyme » ou « pro syn. »
Recommandation 50B
50B. 1. En citant un nomen nudum, on devrait indiquer son statut en ajoutant les mots « nomen nudum » ou « nom.
nud. »
Ex. 1. « Carex bebbii » (Olney, Car. Bor.-Am. 2: 12. 1871), publié sans diagnose ni description, devrait être cité
comme Carex bebbii Olney, nomen nudum (ou nom. nud.).
Recommandation 50C
50C. 1. La citation d’un homonyme postérieur devrait être suivie du nom de l’auteur de l’homonyme antérieur précédé par le mot « non », de préférence avec indication de la date de publication. Dans certains cas, il est à conseiller de citer en outre d’autres homonymes éventuels, précédés du mot « nec ».
Ex. 1. Ulmus racemosa Thomas, Amer. J. Sci. Arts 19 : 170. 1831, non Borkh. 1800 ; Lindera Thunb., Nov. Gen.
Pl. 64. 1783, non Adans. 1763 ; Bartlingia Brongn. in Ann. Sci. Nat. (Paris) 10 : 373. 1827, non Rchb. 1824 nec F.
Muell. 1882.
Recommandation 50D
50D.1. Les erreurs d’interprétation ne devraient pas être insérées dans les synonymies, mais ajoutées à la suite. Tout nom mal appliqué devrait être signalé par l’expression « auct. non », suivie du nom de l’auteur original et des citations bibliographiques concernant l’erreur d’interprétation.
Ex. 1. Ficus stortophylla Warb. in Ann. Mus. Congo Belge, B, Bot. ser. 4, 1: 32 1904. F. irumuënsis De Wild., Pl. Bequaert. 1 : 341. 1922. « F. exasperata » auct. non Vahl : De Wildeman & T. Durand, Ann. Mus. Congo Belge, B, Bot. ser. 2, 1: 54. 1899 ; De Wildeman, Miss. Em. Laurent : 26. 1905 ; T. Durand & H. Durand, Syll. Fl. Congol. : 505. 1909.
Recommandation 50E
50E.1. Si un nom de famille, de genre ou d’espèce, est accepté comme nomen conservandum (voir l’Art. 14 et l’App. II-III), l’abréviation « nom. cons. » devrait être ajouté dans une citation complète.
Ex. 1. Protea L., Mant. Pl. 187. 1771, nom. cons., non L. 1753 ; Combretum Loefl. (1758), nom. cons. [= Grislea L. 1753].
50E.2. Si un nom a été adopté par Fries ou par Persoon, et en conséquence sanctionné (voir l’Art. 13.1 (d) et 7.8), « : Fr. » ou « : Pers. » devrait être ajouté dans la citation complète. La même convention devrait être employée pour le basionyme du nom sanctionné, s’il y en a un, et pour toutes les combinaisons basées indistinctement sur le nom sanctionné ou son basionyme.
Ex. 2. Boletus piperatus Bull. (Herb. France : t. 451, f. 2. 1790) a été accepté dans Fries (Syst. Mycol. 1 : 388. 1821) et a été ainsi sanctionné. On peut donc le citer B. piperatus Bull. : Fr., et la combinaison subséquente basée sur lui, comme Chalciporus piperatus (Bull. : Fr.) Bataille.
Ex. 3. Agaricus sarcocephalus Fr. 1815: Fr. fut sanctioné comme Agaricus compactus [sans rang] sarcocephalus (Fr. : Fr.) Fr. 1821 ; Psathyrella sarcocephala (Fr. : Fr.) Singer est une combinaison subséquente basée sur lui.
Recommandation 50F
50F.1. Si un nom est cité avec des altérations de la forme exacte sous laquelle il a été publié à l’origine, il est bon
d’ajouter cette dernière, de préférence entre guillemets, dans la citation complète.
Ex. 1. Pyrus calleryana Decne. (P. mairei H. Lév. in Repert. Spec. Nov. Regni Veg. 12 : 189. 1913, « Pirus »).
Ex. 2. Zanthoxylum cribrosum Spreng., Syst. Veg. 1 : 946. 1825, « Xanthoxylon ». (Z. caribaeum var. floridanum (Nutt.) A. Gray in Proc. Amer. Acad. Arts 23 : 225. 1888, « Xanthoxylum »).
Ex. 3. Spathiphyllum solomonense Nicolson in Amer. J. Bot. 54 : 496. 1967, « solomonensis ». |
Petite bibliographie sur les plantes carnivores
-
Gérard BLONDEAU, PLANTES CARNIVORES, EDITIONS DE VECCHI, 1996. Très pratique, chaque plante est décrite sous forme de fiches illustrées de photographies. De petits schémas expliquent les principes généraux d'alimentation, de culture, etc.
-
Adrian SLACK, LES PLANTES CARNIVORES, Comment les cultiver?, DIAGONE-CALMANN-LEVY, 1988. Assez complet sur les principes de culture, il reste une référence par un vrai spécialiste mais il n'est pas toujours évident de trouver une info précise sur une espèce particulière, à moins de tout lire sur le genre en question. L'organisation des groupes taxonomiques est décrite en détail et les hybrides sont bien répertoriés.
-
Marcel LECOUFLE, Comment choisir et cultiver vos PLANTES CARNIVORES, éditions BORDAS, 1989. Une variété de plantes assez importante est traitées par une présentation didactique mais il y a quelques erreurs et des bizarreries, comme des modes de reproduction différents pour les divers Sarracenia alors que tous les modes marchent très bien avec toutes les espèces ! Il y a aussi quelques belles photographies ainsi qu'une bonne bibliographie (principalement en anglais).
-
Paul STAROSTA, Jean-Jacques LABAT, L'univers des PLANTES CARNIVORES, Du May, 1993. Vraiment de très belles photographies (Starosta), sur papier glacé et en grand format, qui ne pourront pas vous laisser indifférents.
-
M. BAFFRAY, F. BRICE, Ph. DANTON, J.-P. TOURNIER, Nature et Culture des Plantes Carnivores, EDISUD, 1989. Excellent livre sur l'ensemble des plantes carnivores. La présentation est ordonnée (souvent sous forme de tableaux) et très bien structurée ce qui permet de trouver les informations très facilement. Pour chaque famille, on trouve des renseignements sur les milieux naturels ce qui est plutôt rare. Reste que le dessin représentant des plantules de Dionée en formation sur le piège est pour moi purement imaginaire : depuis presque 20 ans que j'en cultive je n'ai jamais observé, ni entendu parlé de, ni même vu la photo d'une chose pareille... Ce livre ressemble donc plus à une très bonne compilation de sources externes que issue d'un vraie expérience personnelle.
J. Philippe ROSELLO
|